
Синтез белка биология. Синтез белка в клетках мышц
В синтезе белков из аминокислот можно выделить три этапа.
Первый этап – транскрипция – был описан в предыдущей теме. Он состоит в образовании молекул РНК на матрицах ДНК. Для синтеза белка особое значение имеет синтез матричных или информационных РНК, так как здесь записана информация о будущем белке. Транскрипция протекает в ядре клеток. Затем с помощью специальных ферментов, образовавшаяся матричная РНК перемещается в цитоплазму.
Второй этап называется рекогниция. Аминокислоты избирательно связываются с своими переносчиками транспортными РНК .
Все т-РНК построены сходным образом. Молекула каждой т-РНК представляет собой полинуклеотидную цепь, изогнутую в виде «клеверного листа». Молекулы т-РНК устроены таким образом, что имеют разные концы, имеющие сродство и с м-РНК (антикодон) и с аминокислотами. Т-РНК имеет в клетке 60 разновидностей.
Для соединения аминокислот с транспортными РНК служит особый фермент т-РНК синтетаза или, точнее, амино-ацил – т-РНК синтетаза.
Третий этап биосинтеза белка называется трансляция. Он происходит на рибосомах. Каждая рибосома состоит из двух частей – большой и малой субъединиц. Они состоят из рибосомных РНК и белков.
Трансляция начинается с присоединения матричной РНК к рибосоме. Затем к образовавшемуся комплексу начинают присоединяться т-РНК с аминокислотами. Присоединение это происходит путем связывания антикодона т-РНК к кодону информационной РНК на основании принципа комплементарности. Одновременно к рибосоме могут присоединится не более двух т-РНК. Далее аминокислоты соединяются друг с другом пептидной связью, образуя постепенно полипептид. После этого рибосома передвигает информационную РНК ровно на один кодон. Дальше процесс повторяется снова до тех пор пока информационная РНК не закончится. На конце и-РНК находятся бессмысленные кодоны, которые являются точками в записи и одновременно командой для рибосомы, что она должна отделиться от и-РНК
Таким образом, можно выделить несколько особенности биосинтеза белков.
1. Первичная структура белков формируется строго на основе данных записанных в молекулах ДНК и информационных РНК,
2. Высшие структуры белка (вторичная, третичная, четвертичная) возникают самопроизвольно на основе первичной структуры.
3. В некоторых случаях полипептидная цепь после завершения синтеза подвергается незначительной химической модификации, в результате чего в ней появляются некодируемые аминокислоты, не относящиеся к 20 обычным. Примером такого превращения является белок коллаген, где аминокислоты лизин и пролин превращаются в оксипролин и оксилизин.
4. Синтез белков в организме ускоряется соматотропным гормоном и гормоном тестостероном.
5. Синтез белков очень энергоемкий процесс, требующий огромного количества АТФ.
6. Многие антибиотики подавляют трансляцию.
Метаболизм аминокислот.
Аминокислоты могут использоваться для синтеза различных небелковых соединений. Например, из аминокислот синтезируется глюкоза, азотистые основания, небелковая часть гемоглобина – гем, гормоны – адреналин, тироксин и такие важные соединения, как креатин, карнитин, принимающие участие в энергетическом обмене.
Часть аминокислот подвергается распаду до углекислого газа, воды и аммиака.
Распад начинается с реакций общих для большинства аминокислот.
К ним относятся.
1. Декарбоксилирование - отщепление от аминокислот карбоксильной группы в виде углекислого газа.
ПФ (пиридоксальфосфат) – кофермент производное витамина В6.
Так, например, образуется гистамин из аминокислоты гистидина. Гистамин – важное сосудорасширяющее вещество.
2. Дезаминирование - отцепление аминогруппы в виде NH3 . У человека дезаминирование аминокислот идет окислительным путем.
3. Трансаминирование – реакция между аминокислотами и α-кетокислотами. В ходе этой реакции её участники обмениваются функциональными группами.

Трансаминированию подвергаются все аминокислоты. Этот процесс – главное превращение аминокислот в организме, так как у него скорость значительно выше, чем у двух первых описанных реакций.
Трансаминирование выполняет две основные функции.
1. За счет этих реакций одни аминокислоты превращаются в другие. При этом общее количество аминокислот не меняется но меняется общее соотношение между ними в организме. С пищей в организм поступают чужеродные белки, у которых аминокислоты находятся в иных пропорциях. Путем трансаминирования происходит корректировка аминокислотного состава организма.
2. Трансаминирование является составной частью процесса косвенного дезаминирования аминокислот – процесса, с которого начинается распад большинства аминокислот.
Схема косвенного дезаминирования.

В результате трансаминирования образуются α-кетокислоты и аммиак. Первые разрушаются до углекислого газа и воды. Аммиак для организма высокотоксичен. Поэтому в организме существуют молекулярные механизмы его обезвреживания.
Биосинтез белка.
Пластический обмен (ассимиляция или анаболизм) – совокупность реакций биологического синтеза. Название этого вида обмена отражает его сущность: из веществ, поступающих в клетку из вне, образуются вещества, подобные веществам клетки.
Рассмотрим одну из важнейших форм пластического обмена – биосинтез белков. Биосинтез белков осуществляется во всех клетках про -и эукариот. Информация о первичной структуре (порядке аминокислот) белковой молекулы закодирована последовательностью нуклеотидов в соответствующем участке молекулы ДНК - гене.
Ген- это участок молекулы ДНК, определяющий порядок аминокислот в молекуле белка. Следовательно, от порядка нуклеотидов в гене зависит порядок аминокислот в полипептиде, т.е. его первичная структура, от которой в свою очередь зависят все другие структуры, свойства и функции белковой молекулы.
Система записи генетической информации в ДНК (и - РНК) в виде определенной последовательности нуклеотидов называется генетическим кодом. Т.е. единица генетического кода (кодон) - это триплет нуклеотидов в ДНК или РНК, кодирующий одну аминокислоту.
Всего генетический код включает 64 кодона, из них 61 кодирующий и 3 некодирующих (кодоны-терминаторы, свидетельствующие об окончании процесса трансляции).
Кодоны-терминаторы в и - РНК: УАА, УАГ, УГА, в ДНК: АТТ, АТЦ, АЦТ.
Начало процесса трансляции определяет кодон-инициатор (АУГ, в ДНК - ТАЦ), кодирующий аминокислоту метионин. Этот кодон первым входит в рибосому. Впоследствии метионин, если он не предусмотрен в качестве первой аминокислоты данного белка, отщепляется.
Генетический код обладает характерными свойствами.
1. Универсальность - код одинаков для всех организмов. Один и тот же триплет (кодон) в любом организме кодирует одну и ту же аминокислоту.
2. Специфичность - каждый кодон шифрует только одну аминокислоту.
3. Вырожденность - большинство аминокислот могут кодироваться несколькими кодонами. Исключение составляют 2 аминокислоты - метионин и триптофан, имеющие лишь по одному варианту кодона.
4. Между генами имеются «знаки препинания» - три специальных триплета (УАА, УАГ, УГА), каждый из которых обозначает прекращение синтеза полипептидной цепи.
5. Внутри гена «знаков препинания» нет.
Для того, чтобы синтезировался белок, информация о последовательности нуклеотидов в его первичной структуре должна быть доставлена к рибосомам. Этот процесс включает два этапа – транскрипцию и трансляцию.
Транскрипция (переписывание) информации происходит путем синтеза на одной из цепей молекулы ДНК одноцепочной молекулы РНК, последовательность нуклеотидов которой точно соответствует последовательности нуклеотидов матрицы – полинуклеотидной цепи ДНК.
Она (и - РНК) является посредником, передающим информацию от ДНК к месту сборки молекул белка в рибосоме. Синтез и - РНК (транскрипция) происходит следующим образом. Фермент (РНК - полимераза) расщепляет двойную цепочку ДНК, и на одной из ее цепей (кодирующей) по принципу комплементарности выстраиваются нуклеотиды РНК. Синтезированная таким образом (матричный синтез) молекула и - РНК выходит в цитоплазму, и на один ее конец нанизываются малые субъединицы рибосом.
Второй этап в биосинтезе белка - трансляция - это перевод последовательности нуклеотидов в молекуле и - РНК в последовательность аминокислот в полипептиде. У прокариот, не имеющих оформленного ядра, рибосомы могут связываться с вновь синтезированной молекулой и - РНК сразу же после ее отделения от ДНК или даже до полного завершения ее синтеза. У эукариот и - РНК сначала должна быть доставлена через ядерную оболочку в цитоплазму. Перенос осуществляется специальными белками, которые образуют комплекс с молекулой и - РНК. Кроме функций переноса эти белки защищают и - РНК от повреждающего действия цитоплазматических ферментов.
В цитоплазме на один из концов и - РНК (а именно на тот, с которого начинается синтез молекулы в ядре) вступает рибосома и начинается синтез полипептида. По мере продвижения по молекуле РНК рибосома транслирует триплет за триплетом, последовательно присоединяя аминокислоты к растущему концу полипептидной цепи. Точное соответствие аминокислоты коду триплета и - РНК обеспечивается т - РНК.
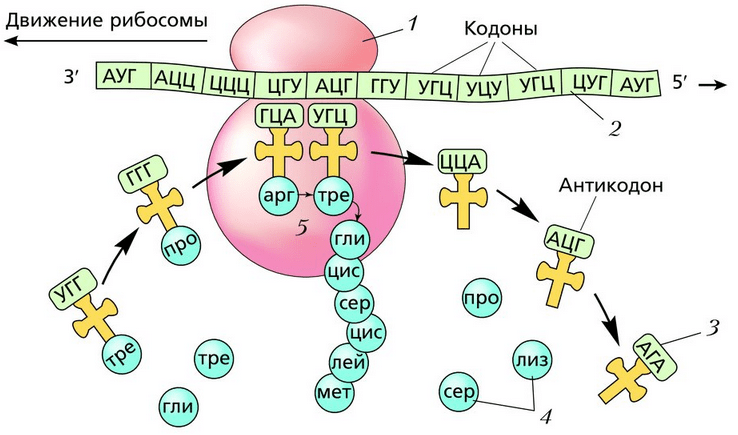
Транспортные РНК (т - РНК) «приносят» аминокислоты в большую субъединицу рибосомы. Молекула т - РНК имеет сложную конфигурацию. На некоторых участках ее между комплементарными нуклеотидами образуются водородные связи, и молекула по форме напоминает лист клевера. На ее верхушке расположен триплет свободных нуклеотидов (антикодон), который соответствует определенной аминокислоте, а основание служит местом прикрепления этой аминокислоты (рис. 1).
Рис. 1. Схема строения транспортной РНК: 1 - водородные связи; 2 - антикодон; 3 -место прикрепления аминокислоты.
Каждая т - РНК может переносить только свою аминокислоту. Т-РНК активируется специальными ферментами, присоединяет свою аминокислоту и транспортирует ее в рибосому. Внутри рибосомы в каждый данный момент находится всего два кодона и-РНК. Если антикодон т-РНК является комплементарным кодону и-РНК, то происходит временное присоединение т-РНК с аминокислотой к и-РНК. Ко второму кодону присоединяется вторая т-РНК, несущая свою аминокислоту. Аминокислоты располагаются рядом в большой субъединице рибосомы, и с помощью ферментов между ними устанавливается пептидная связь. Одновременно разрушается связь между первой аминокислотой и ее т-РНК, и т-РНК уходит из рибосомы за следующей аминокислотой. Рибосома перемещается на один триплет, и процесс повторяется. Так постепенно наращивается молекула полипептида, в которой аминокислоты располагаются в строгом соответствии с порядком кодирующих их триплетов (матричный синтез) (рис. 2).

Рис. 2. Схема бисинтеза белка: 1 - и-РНК; 2 - субъединицы рибосомы; 3 - т-РНК с аминокислотами; 4 - т-РНК без аминокислот; 5 - полипептид; 6 - кодон и-РНК; 7- антикодон т-РНК.
Одна рибосома способна синтезировать полную полипептидную цепь. Однако, нередко по одной молекуле и-РНК движется несколько рибосом. Такие комплексы называются полирибосомами. После завершения синтеза полипептидная цепочка отделяется от матрицы – молекулы и-РНК, сворачивается в спираль и приобретает свойственную ей (вторичную, третичную или четвертичную) структуру. Рибосомы работают очень эффективно: в течение 1с бактериальная рибосома образует полипептидную цепь из 20 аминокислот.
Биосинтез белка и генетический код
Определение 1
Биосинтез белка – ферментативный процесс синтеза белков в клетке. В нём участвуют три структурные элемента клетки – ядро, цитоплазма, рибосомы.
В ядре клетки в молекулах ДНК сохраняется информация о всех белках, которые в ней синтезируются, зашифрованная с помощью четырёхбуквенного кода.
Определение 2
Генетический код – это последовательность расположения нуклеотидов в молекуле ДНК, которая определяет последовательность аминокислот в молекуле белка.
Свойства генетического кода таковы:
Генетический код триплетный, то есть каждой аминокислоте соответствует свой кодовый триплет (кодон ), состоящий из трёх расположенных рядом нуклеотидов.
Пример 1
Аминокислота цистеин кодируется триплетом А-Ц-А, валин – триплетом Ц-А-А.
Код не перекрывается, то есть нуклеотид не может входить в состав двух соседних триплетов.
Код вырожден, то есть одна аминокислота может кодироваться несколькими триплетами.
Пример 2
Аминокислота тирозин кодируется двумя триплетами.
Код не имеет запятых (разделительных знаков), считывание информации происходит тройками нуклеотидов.
Определение 3
Ген – участок молекулы ДНК, который характеризуется определённой последовательностью нуклеотидов и определяет синтез одногой полипептидной цепи.
Код является универсальным, то есть единым для всех живых организмов – от бактерий до человека. У всех организмов есть одни и те же 20 аминокислот, которые кодируются одними и теми же триплетами.
Этапы биосинтеза белка: транскрипция и трансляция
Структура любой белковой молекулы закодирована в ДНК, которая не участвует непосредственно в её синтезе. Она служит лишь матрицей для синтеза РНК.
Процесс биосинтеза белка происходит на рибосомах, которые расположены преимущественно в цитоплазме. Значит, для осуществления передачи к месту синтеза белка генетической информации из ДНК нужен посредник. Эту функцию выполняет иРНК.
Определение 4
Процесс синтеза молекулы иРНК на одной цепи молекулы ДНК на основании принципа комплементарности называется транскрипцией , или переписыванием.
Транскрипция происходит в ядре клетки.
Процесс транскрипции осуществляется одновременно не на всей молекуле ДНК, а лишь на её небольшом участке, который отвечает определённому гену. При этом происходит раскручивание части двойной спирали ДНК и короткий участок одной из цепей оголяется – теперь он будет выполнять роль матрицы для синтеза иРНК.
Потом вдоль этой цепи двигается фермент РНК-полимераза, соединяющий нуклеотиды в цепь иРНК, которая удлиняется.
Замечание 2
Транскрипция может одновременно происходить и на нескольких генах одной хромосомы и на генах разных хромосомах.
Образованная в результате иРНК содержит последовательность нуклеотидов, которая является точной копией последовательности нуклеотидов на матрице.
Замечание 3
Если в молекуле ДНК есть азотистое основание цитозин, то в иРНК – гуанин и наоборот. Комплементарной парой в ДНК является аденин – тимин, а РНК вместо тимина содержит урацил.
На специальных генах синтезируются и два другие типа РНК – тРНК и рРНК.
Начало и окончание синтеза всех типов РНК на матрице ДНК строго фиксированы специальными триплетами, которые контролируют запуск (инициирующие) и остановку (терминальные) синтеза. Они выполняют функции «разделительных знаков» между генами.
Соединение тРНК с аминокислотами происходит в цитоплазме. Молекула тРНК формой напоминает листик клевера, на его верхушке расположен антикодон – триплет нуклеотидов, который кодирует аминокислоту, которую переносит данная тРНК.
Сколько видов аминокислот, столько существует и тРНК.
Замечание 4
Поскольку много аминокислот могут кодироваться несколькими триплетами, то количество тРНК больше 20 (известно около 60 тРНК).
Соединение тРНК с аминокислотами происходит с участием ферментов. Молекулы тРНК транспортируют аминокислоты к рибосомам.
Определение 5
Трансляция – это процесс, в результате которого информация о структуре белка, записанная в иРНК в виде последовательности нуклеотидов, реализуется в виде последовательности аминокислот в молекуле белка, которая синтезируется.
Этот процесс осуществляется в рибосомах.
Сначала иРНК присоединяется к рибосоме. На иРНК «нанизывается» первая рибосома, которая синтезирует белок. По мере продвижения рибосомы на конец иРНК, который освободился, «нанизывается» новая рибосома. На одной иРНК могут находиться одновременно более 80 рибосом, которые синтезируют один и тот же белок. Такая группа рибосом, соединённых с одной иРНК, называется полирибосомой , или полисомой . Вид белка, который синтезируется, определяется не рибосомой, а информацией, записанной на иРНК. Одна и та же рибосома способна синтезировать разные белки. После завершения синтеза белка рибосома отделяется от иРНК, а белок поступает в эндоплазматическую сеть.
Каждая рибосома состоит из двух субъединиц – малой и большой. Молекула иРНК присоединяется к малой субъединице. В месте контакта рибосомы и иРН находятся 6 нуклеотидов (2 триплета). К одному из них всё время подходят из цитоплазмы тРНК с разными аминокислотами и касаются антикодоном кодона иРНК. Если триплеты кодона и антикодона оказываются комплементарными, между аминокислотой уже синтезированной части белка и аминокислотой, которая доставляется тРНК, возникает пептидная связь. Соединение аминокислот в молекулу белка осуществляется с участием фермента синтетазы. Молекула тРНК отдаёт аминокислоту и переходит в цитоплазму, а рибосома передвигается на один триплет нуклеотидов. Так последовательно синтезируется полипептидная цепь. Продолжается всё это до тех пор, пока рибосома не дойдёт к одному из трёх терминирующих кодонов: УАА, УАГ или УГА. После этого синтез белка прекращается.
Замечание 5
Таким образом, последовательность кодонов иРНК определяет последовательность включения аминокислот в цепь белка. Синтезированные белки поступают в каналы эндоплазматического ретикулюма. Одна молекула белка в клетке синтезируется за 1 - 2 минуты.
Сначала, установите последовательность этапов биосинтеза белка, начиная с транскрипции. Всю последовательность процессов, происходящих при синтезе белковых молекул, можно объединить в 2 этапа:
Транскрипция.
Трансляция.
Структурными единицами наследственной информации являются гены – участки молекулы ДНК, кодирующие синтез определенного белка. По химической организации материал наследственности и изменчивости про- и эукариот принципиально не отличается. Генетический материал в них представлен в молекуле ДНК, общим является также принцип записи наследственной информации и генетический код. Одни и те же аминокислоты у про — и эукариот шифруются одинаковыми кодонами.
Геном современных прокариотических клеток характеризуется относительно небольшими размерами, ДНК кишечной палочки имеет вид кольца, длиной около 1 мм. Она содержит 4 х 10 6 пар нуклеотидов, образующих около 4000 генов. В 1961 г. Ф. Жакоб и Ж. Моно открыли цистронную, или непрерывную организацию генов прокариот, которые полностью состоят из кодирующих нуклеотидных последовательностей, и они целиком реализуются в ходе синтеза белков. Наследственный материал молекулы ДНК прокариот располагается непосредственно в цитоплазме клетки, где также находятся необходимые для экспрессии генов тРНК и ферменты.Экспрессия- это функциональная активность генов, или выражение генов. Поэтому синтезированная с ДНК иРНК способна сразу выполнять функцию матрицы в процессе трансляции синтеза белка.
Геном эукариот содержит значительно больше наследственного материала. У человека общая длина ДНК в диплоидном наборе хромосом составляет около 174 см. Она содержит 3 х 10 9 пар нуклеотидов и включает до 100000 генов. В 1977 г. была обнаружена прерывистость в строении большинства генов эукариот, получивший название «мозаичный» ген. Для него характерны кодирующие нуклеотидные последовательности экзонные и интронные участки. Для синтеза белка используется только информация экзонов. Количество интронов варьирует в разных генах. Установлено,что ген овальбумина кур включает 7 интронов, а ген проколлагена млекопитающих – 50. Функции молчащей ДНК – интронов окончательно не выяснены. Предполагают, что они обеспечивают: 1) структурную организацию хроматина; 2) некоторые из них, очевидно, участвуют в регуляции экспрессии генов; 3) интроны можно считать запасом информации для изменчивости; 4) они могут играть защитную роль, принимая на себя действие мутагенов.
Транскрипция
Процесс переписывания информации в ядре клетки с участка молекулы ДНК на молекулу мРНК (иРНК) называется транскрипция (лат. Transcriptio – переписывание). Синтезируется первичный продукт гена- мРНК. Это первый этап белкового синтеза. На соответствующем участке ДНК фермент РНК–полимераза узнает знак начала транскрипции – промотр. Стартовой точкой считается первый нуклеотид ДНК, который включается ферментом в РНК-транскрипт. Как правило, кодирующие участки начинаются кодоном АУГ, иногда у бактерий используется ГУГ. Когда РНК-полимераза связывается с промотором, происходит локальное расплетание двойной спирали ДНК и копируется одна из цепей по принципу комплементарности. Синтезируется мРНК, скорость сборки её достигает 50 нуклеотидов в секунду. По мере движения РНК–полимеразы, растёт цепь мРНК, и когда фермент достигнет конца копирующего участка – терминатора , мРНК отходит от матрицы. Двойная спираль ДНК позади фермента восстанавливается.
Транскипция прокариот осуществляется в цитоплазме. В связи с тем, что ДНК целиком состоит из кодирующих нуклеотидных последовательностей, поэтому синтезированная мРНК сразу выполняет функцию матрицы для трансляции (см. выше).
Транскрипция мРНК у эукариот происходит в ядре. Она начинается синтезом больших по размерам молекул — предшественников (про-мРНК), называемых незрелой, или ядерной РНК.Первичный продукт гена- про-мРНК является точной копией транскрибированного участка ДНК, включает экзоны и интроны. Процесс формирования зрелых молекул РНК из предшественников называется процессингом . Созревание мРНК происходит путём сплайсинга – это вырезания ферментами рестриктаз интронов и соединение участков с транскрибируемыми последовательностями экзонов ферментами лигаз. (Рис.).Зрелая мРНК значительно короче молекул-предшественников про – мРНК, размеры интронов в них варьирует от 100 до 1000 нуклеотидов и более. На долю интронов приходится около 80% всей незрелой мРНК.
В настоящее время доказана возможность альтернативного сплайсинга, при котором из одного первичного транскрипта могут удалятся в разных его участках нуклеотидные последовательности и будут образовываться несколько зрелых мРНК. Данный вид сплайсинга характерен в системе генов иммуноглобулинов у млекопитающих, что даёт возможность формировать на основе одного транскрипта мРНК разные виды антител.
По завершению процессинга зрелая мРНК проходит отбор перед выходом из ядра. Установлено, что в цитоплазму попадает всего 5% зрелой мРНК, а остальная часть расщепляется в ядре.
Трансляция
Трансляция (лат. Translatio — передача, перенесение) — перевод информации, заключенной в последовательности нуклеотидов молекулы мРНК,в последовательность аминокислот полипептидной цепи (Рис. 10). Это второй этап белкового синтеза. Перенос зрелой мРНК через поры ядерной оболочки производят специальные белки, которые образуют комплекс с молекулой РНК. Кроме транспорта мРНК, эти белки защищают мРНК от повреждающего действия цитоплазматических ферментов. В процессе трансляции центральная роль принадлежит тРНК, они обеспечивают точное соответствие аминокислоты коду триплета мРНК. Процесс трансляции- декодирования происходит в рибосомах и осуществляется в направлении от 5 к 3 , Комплекс мРНК и рибосом называется полисомой.
В ходе трансляции можно выделить три фазы: инициацию, элонгацию и терминацию.
Инициация.
На этом этапе происходит сборка всего комплекса, участвующего в синтезе молекулы белка. Происходит объединение двух субъединиц рибосом на определённом участке мРНК, присоединение к ней первой аминоацил – тРНК и этим задаётся рамка считывания информации. В молекуле любой м-РНК есть участок, комплементарный р-РНК малой субединицы рибосомы и специфически ею управляемый. Рядом с ним находится инициирующий стартовый кодон АУГ, который кодирует аминокислоту метионин.Фаза инициации завершается образованием комплекса:рибосома, -мРНК- инициирующая аминоацил-тРНК.
Элонгация
— она включает все реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. На рибосоме имеется два участка для связывания двух молекул т-РНК. В одном участке-пептидильном(П) находится первая т-РНК с аминокислотой метионин и с него начинается синтез любой молекулы белка. Во второй участок рибосомы- аминоацильный (А) поступает вторая молекула т-РНК и присоединяется к своему кодону. Между метионином и второй аминокислотой образуется пептидная связь. Вторая т-РНК перемещается вместе со своим кодоном м-РНК в пептидильный центр. Перемещение т-РНК с полипептидной цепочкой из аминоацильного центра в пептидильный сопровождается продвижением рибосомы по м-РНК на шаг, соответствующий одному кодону. Т-РНК, доставившая метионин, возвращается в цитоплазму, амноацильный центр освобождается. В него поступает новая т-РНК с аминокислотой, зашифрованной очередным кодоном. Между третьей и второй аминокислотами образуется пептидная связь и третья т-РНК вместе с кодоном м-РНК перемещается в пептидильный центр.Процесс элонгации, удлинения белковой цепи. Продолжается до тех пор, пока в рибосому не попадёт один из трёх кодонов, не кодирующих аминокислоты. Это кодон — терминатор и для него не существует соответствущей т-РНК, поэтому ни одна из т-РНК не может занять место в аминоацильном центре.
Терминация
– завершение синтеза полипептида. Она связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ, УГА), когда он будет входить в аминоацильный центр. К рибосоме присоединяется специальный фактор терминации, который способствует разъединению субъединиц рибосомы и освобождению синтезированной молекулы белка. К последней аминокислоте пептида присоединяется вода и её карбоксильный конец отделяется от т-РНК.
Сборка пептидной цепи осуществляется с большой скоростью. У бактерий при температуре 37°С она выражается в добавлении к полипептиду от 12 до 17 аминокислот в секунду. В эукариотических клетках к полипептиду добавляется две аминокислоты в одну секунду.
Синтезированная полипептидная цепь затем поступает в комплекс Гольджи, где завершается построение белковой молекулы (последовательно возникают вторая, третья, четвертая структуры). Здесь же происходит комплексование белковых молекул с жирами и углеводами.
Весь процесс биосинтеза белка представлен в виде схемы: ДНК ® про иРНК ® мРНК ® полипептидная цепь ® белок® комплексование белков и их преобразование в функционально активные молекулы.
Этапы реализации наследственной информации также протекают сходным образом: сначала она транскрибируется в нуклеотидную последовательность мРНК, а затем транслируется в аминокислотную последовательность полипептида на рибосомах с участием тРНК.
Транскрипция эукариот осуществляется под действием трех ядерных РНК-полимераз. РНК-полимераза 1 находится в ядрышках и отвечает за транскрипцию генов рРНК. РНК-полимераза 2 находится в ядерном соке и отвечает за синтез предшественника мРНК. РНК-полимераза 3 –небольшая фракция в ядерном соке, которая осуществляет синтез малых рРНК и тРНК. РНК-полимеразы специфически узнают нуклеотидную последовательность транскрипции-промотор. Эукариотическая мРНК вначале синтезируется в виде предшественницы (про- иРНК), на нее списывается информация с экзонов и интронов. Синтезированная мРНК обладает большими, чем необходимо для трансляции размерами и оказывается менее стабильной.
В процессе созревания молекулы мРНК с помощью ферментов рестриктаз вырезаются интроны, а с помощью ферментов – лигаз сшиваются экзоны. Созревание мРНК называется процессингом, сшивание экзонов называется сплайсингом. Таким образом, зрелая мРНК содержит только экзоны и она значительно короче её предшественницы – про- иРНК. Размеры интронов варьируют от 100 до 10000 нуклеотидов и более. На долю интонов приходится около 80% всей незрелой мРНК. В настоящее время доказана возможность альтернативного сплайсинга, при котором из одного первичного транскрипта могут удаляться в разных его участках нуклеотидные последовательности и будут образовываться несколько зрелых мРНК. Данный вид сплайсинга характерен в системе генов иммуноглобулинов у млекопитающих, что даёт возможность формировать на основе одного транскрипта мРНК разные виды антител. По завершению процессинга зрелая мРНК проходит отбор перед выходом в цитоплазму из ядра. Установлено, что попадает всего 5% зрелой мРНК, а остальная часть расщепляется в ядре. Преобразование первичных транскриптонов эукариотических генов, связанное с их экзон-интронной организацией, и в связи с переходом зрелой мРНК из ядра в цитоплазму, определяет особенности реализации генетической информации эукариот. Следовательно, мозаичный ген эукариот не является геном цистроном, так как не вся последовательность ДНК используется для синтеза белка.
Процесс белкового биосинтеза чрезвычайно важен для клетки. Поскольку белки являются сложными веществами, которые играют основную роль в тканях, они незаменимы. По этой причине в клетке реализована целая цепь процессов белкового биосинтеза, которая протекает в нескольких органеллах. Это гарантирует клетке воспроизведение и возможность существования.
Сущность процесса биосинтеза белка
Единственное место синтеза белков - это шероховатая Здесь располагается основная масса рибосом, которые ответственны за образование полипептидной цепочки. Однако до того как начнется этап трансляции (процесс синтеза белка), требуется активация гена, в котором хранится информация о белковой структуре. После этого требуется копирование данного участка ДНК (или РНК, если рассматривается бактериальный биосинтез).
После копирования ДНК требуется процесс создания информационной РНК. На ее основании будет выполняться синтез белковой цепочки. Причем все этапы, которые протекают с вовлечением нуклеиновых кислот, должны происходить в Однако это не место, где происходит синтез белка. где осуществляется подготовка к биосинтезу.
Рибосомальный биосинтез белка
Основное место, где происходит синтез белка, - клеточная органелла, состоящая из двух субъединиц. Таких структур в клетке огромное количество, и они в основном расположены на мембранах шероховатой эндоплазматической сети. Сам биосинтез происходит так: образованная в ядре клетки информационная РНК выходит сквозь нуклеарные поры в цитоплазму и встречается с рибосомой. Затем иРНК проталкивается в промежуток между субъединицами рибосомы, после чего происходит фиксация первой аминокислоты.
К месту, где происходит синтез белка, аминокислоты подаются при помощи Одна такая молекула может однократно приносить по одной аминокислоте. Они присоединяются по очереди в зависимости от последовательности кодонов информационной РНК. Также синтез может прекращаться на некоторое время.
При продвижении по иРНК рибосома может попадать на участки (интроны), которые не кодируют аминокислоты. В этих местах рибосома просто продвигается по иРНК, но присоединения аминокислот к цепочке не происходит. Как только рибосома достигает экзона, то есть участка, который кодирует кислоту, тогда она снова присоединяется к полипептиду.
Постсинтетическая модификация белков
После достижения рибосомой стоп-кодона информационной РНК процесс непосредственного синтеза завершается. Однако полученная молекула имеет первичную структуру и пока не может выполнять зарезервированных для нее функций. Для того чтобы полноценно функционировать, молекула должна организоваться в определенную структуру: вторичную, третичную или еще более сложную - четвертичную.

Структурная организация белка
Вторичная структура - первая стадия структурной организации. Для ее достижения первичная полипептидная цепочка должна спирализоваться (образовать альфа-спирали) или загибаться (создать бета-слои). Затем, для того чтобы занимать еще меньше места по длине, молекула еще больше стягивается и сматывается в клубок за счет водородных, ковалентных и ионных связей, а также межатомных взаимодействий. Таким образом, получается глобулярная

Четвертичная белковая структура
Четвертичная структура самая сложная из всех. Она состоит из нескольких участков с глобулярным строением, соединенных фибриллярными нитями полипептида. Вдобавок третичная и четвертичная структура могут содержать углеводный или липидный остаток, что расширяет спектр функций белка. В частности, гликопротеиды, белка и углевода, являются иммуноглобулинами и выполняют защитную функцию. Также гликопротеиды располагаются на мембранах клеток и работают рецепторами. Однако модифицируется молекула не там, где происходит синтез белка, а в гладкой эндоплазматической сети. Здесь существует возможность присоединения липидов, металлов и углеводов к доменам белков.